

Cellular senescence has moved from a curiosity of cell culture into one of the most actively investigated processes in biomedicine. In fact, the longevity and anti-senescence market is expected to reach 44.2 billion by 2030 [13].
First described as the irreversible replicative arrest of cultured fibroblasts, senescence is now understood as a dynamic, highly regulated stress response that shapes embryonic development, wound repair, aging, and tumor biology alike [1, 2]. What makes senescence such a compelling and frustrating research target is its dual nature: the same cellular program that protects tissues from damaged or pre-malignant cells can also become a chronic source of tissue dysfunction when senescent cells accumulate and persist. [2, 3]. Untangling these opposing roles requires robust, reproducible cell-based assays, and that is exactly where modern cytokine and cell-based assay platforms are proving indispensable.
What Cellular Senescence Looks Like Under the Microscope
Senescent cells exit the cell cycle permanently in response to stressors such as telomere attrition, DNA damage, oncogene activation, or epigenetic disruption, yet they remain metabolically active and acquire a distinctive secretory program [3]. This program, known as the senescence-associated secretory phenotype (SASP), consists of pro-inflammatory cytokines, chemokines, growth factors, and matrix-remodeling proteases that senescent cells release into their microenvironment. [3, 4]. The SASP is the mechanistic thread connecting senescence to aging, regeneration, and cancer because it is largely through these secreted factors that senescent cells communicate with neighboring tissue and influence its fate. [4].
No single biomarker reliably identifies a senescent cell, which is why researchers typically combine several assay types [2, 3]:
- Senescence-associated β-galactosidase (SA-β-Gal) staining, the classic histochemical readout of lysosomal β-galactosidase activity at pH 6.0, still the most widely used screening assay despite known limitations around confluency-related false positives [3].
- Cell-cycle arrest markers, including p16 INK4a, p21 CIP1, and p53 expression, measured by western blot, qPCR, or immunofluorescence [2, 3].
- DNA damage markers such as γ-H2AX foci, which flag the persistent DNA damage response that frequently underlies senescence induction [3].
- SASP profiling, using immunoassays such as ELISA, multiplex, or Luminex-based panels to quantify cytokines like IL-6, IL-8, and TNF-α directly from conditioned media [2, 3].
- Proliferation and cell-cycle assays, including BrdU/EdU incorporation, which distinguish true senescent arrest from reversible quiescence [3].
At SBH Sciences, senescence is assessed using this exact multi-marker framework: SA-β-galactosidase activity, p16 and p21 expression, and functional secretion of IL-6 as a direct SASP readout. Critically, senescence is induced in experimental cell models using three well-characterized stressors: the genotoxic anthracycline doxorubicin, hydrogen peroxide-driven oxidative stress, and G3-YSD. This covers DNA damage, oxidative, and additional stress-induced senescence pathways respectively. This diversity of induction methods matters because SASP composition can vary by senescence trigger, and assays validated across multiple induction models are more likely to reflect the heterogeneity seen in human tissues and tumors.
Because senescent cells are defined more by a constellation of features than by a single marker, cytokine and growth-factor bioassays that quantify the SASP functionally, rather than solely through gene expression, have become an essential complement to histochemical staining. [1, 2].
Cytokine Bioassays: Reading the SASP as a Functional Signal
This is where cell-based cytokine bioassays earn their place in the senescence toolkit. Rather than simply detecting that a cytokine is present, a well-designed bioassay measures whether that cytokine is biologically active by determining whether it actually triggers the downstream proliferative or signaling response in a responsive cell line. This functional readout matters enormously for senescence research, where the SASP's effects on surrounding cells, not just its composition, determine whether senescence ends up suppressing or promoting disease [3, 4].
As an illustration, the chart below shows a representative dose-response bioassay developed by SBH Sciences for human IL-6, one of the most consistently elevated cytokines in the SASP and a central signaling node in senescence-associated inflammation [4, 5]. The assay uses the IL-6-dependent 7TD1 cell line, which proliferates in direct proportion to IL-6 concentration; proliferation is read out colorimetrically (OD 490 nm) and fit to a sigmoidal dose-response curve spanning roughly three orders of magnitude of IL-6 concentration (0.001–1.0 ng/mL).
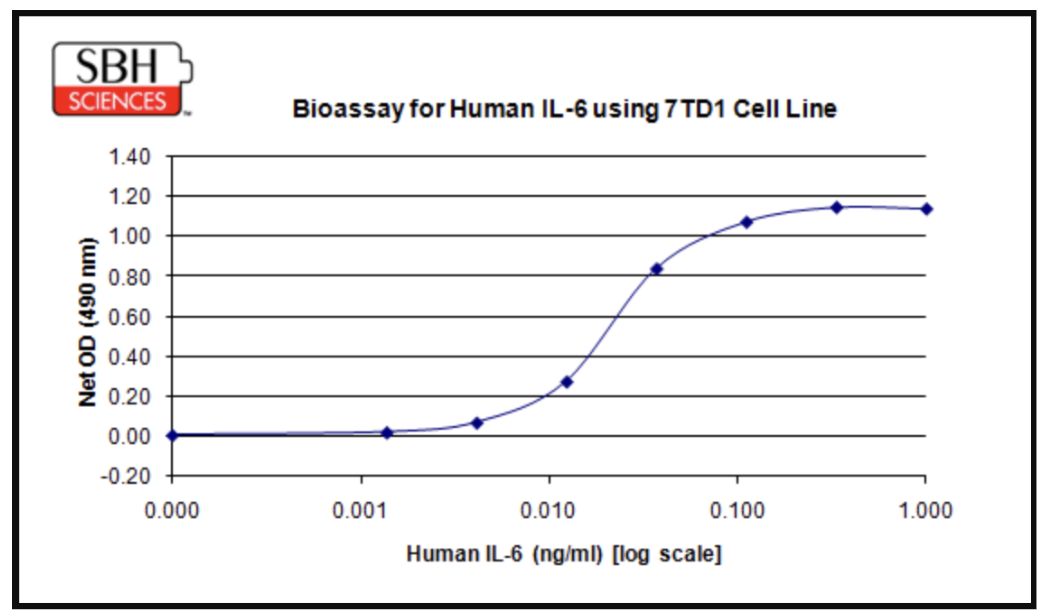
The resulting EC50 and the shape of the curve give researchers a quantitative, reproducible way to compare IL-6 bioactivity across conditions. For example, they can contrast conditioned media from senescent and proliferating cell cultures or test whether a candidate senomorphic or anti-inflammatory compound dampens IL-6 signaling output rather than simply lowering protein concentration.
This kind of assay framework generalizes well beyond IL-6. SASP biology is built on a relatively conserved cytokine and growth factor signature (IL-6, IL-8, IL-1α/β, TNF-α, GM-CSF, VEGF, and a range of matrix metalloproteinases recur across senescence models [3, 4]) and validated bioassays exist for these, alongside neutralization formats that test whether blocking a given cytokine reverses a senescence-associated phenotype. Used together, a panel of such assays lets researchers move past a static inventory of "what's in the SASP" toward a functional picture of what the SASP is actually doing to the surrounding tissue.
Senescence in Regenerative Medicine: A Double-Edged Signal
Senescence was once framed almost exclusively as a hallmark of aging to be eliminated. That view has been substantially revised. Programmed senescence occurs transiently during embryonic development and tissue repair, and a growing body of work shows it is an active, beneficial participant in regeneration [6, 7]. In zebrafish fin regeneration, removing senescent cells at the wound site impairs recovery, establishing senescence as a functional driver rather than a bystander [6]. Comparable biology holds in mammals: senescent synovial fibroblasts transiently accumulate during rat meniscus repair, and a transient wave of pro-reparative senescent fibroblasts supports wound closure in young skin, while its dysregulation contributes to delayed healing in aged tissue [7].
It all comes down to when these changes occur and how the body handles their removal. A short burst of senescence with efficient immune-mediated clearance appears regenerative; a chronic, unresolved burden is degenerative [6, 7]. Cytokine bioassays that quantify SASP factors such as PDGF-AA, IL-6, and matrix-remodeling proteins across a repair time course give researchers a way to characterize that window experimentally. Coupling SA-β-Gal and p16/p21 staining with functional cytokine output assays makes it possible to ask not just whether senescent cells are present, but whether their secretory profile is pro-regenerative or pro-fibrotic — a distinction with direct relevance to stem cell therapy, wound-healing formulations, and tissue engineering [3].
Senescence Models in Cancer Research: Suppressor, Saboteur, or Both?
Senescence's relationship with cancer is more paradoxical than its role in regeneration. Oncogene-induced senescence acts as a bona fide tumor-suppressive barrier: cells experiencing aberrant RAS, BRAF, or other oncogenic signaling can enter stable, irreversible arrest rather than progress toward transformation, and cancer cells can be pushed into this state therapeutically through radiation, chemotherapy, or CDK4/6 inhibition [8, 9].
The other side is more troubling. Senescent cells embedded within or around a tumor secrete a SASP that actively remodels the microenvironment toward disease progression. IL-6 and IL-8 increase breast cancer cell invasiveness and drive epithelial-to-mesenchymal transition (EMT), while IL-6 signaling through STAT3 and NF-κB reprograms non-stem tumor cells toward a stem-like, more aggressive state in a self-amplifying loop [4, 9]. SASP-rich microenvironments further facilitate immune evasion by recruiting myeloid-derived suppressor cells and impairing clearance of both senescent and malignant cells [4]. The very secretory program that aids wound healing can, in a tumor context, become a driver of cancer cell plasticity, drug resistance, and relapse [4, 9, 12].
This dual identity is why senescence has become an active frontier for drug discovery on two distinct fronts simultaneously:
Pro-senescence approaches aim to push cancer cells into a stable arrest as a therapeutic endpoint, often as the "first punch" of a sequential treatment strategy [9]. Validating that a candidate compound truly induces senescence, rather than simple cytotoxicity or transient growth arrest, requires the same multi-marker assay logic used in aging research: SA-β-Gal staining, p16/p21 induction, and morphology-based profiling, increasingly supported by machine-learning classifiers built to distinguish senescence from toxicity and quiescence in high-throughput screens [10].
Senolytic and senomorphic approaches take the opposite tack, either selectively eliminating senescent cells (senolytics) or suppressing their inflammatory output without killing them (senomorphics), with the goal of clearing the harmful SASP-driven microenvironment that fuels tumor progression and treatment resistance [9, 11]. Both strategies depend on functional cytokine assays to confirm target engagement and ask whether the candidate compound actually reduces IL-6, IL-8, or TNF-α bioactivity in conditioned media, and if that reduction tracks with reduced invasiveness or stemness markers in co-culture models [4, 11].
The scale of industry investment underscores how mature this space is becoming. AstraZeneca, for example, has progressed navitoclax, a BCL-2/BCL-xL inhibitor with well-documented senolytic activity, through multiple combination trials across myelofibrosis, small-cell lung cancer, and solid tumor indications, while continuing to build a broader senescence-targeting portfolio [9]. The fact that a single major pharmaceutical company can now field several senescence-related clinical programs simultaneously reflects both the mechanistic coherence of the field and the growing confidence that functional assay evidence from preclinical models translates meaningfully to patient stratification strategies.
Experimental rigor demands well-validated positive controls. SBH Sciences employs standard control panels in all senescence experiments: navitoclax (ABT-263) at 0.5 μM as a senolytic benchmark, metformin at 1 mM for mTOR-linked senomorphic activity, and rapamycin at 100 nM as a canonical SASP suppressor via mTORC1 inhibition [9, 11]. Anchoring screens to compounds with established clinical and mechanistic pedigree, including navitoclax, the same asset in AstraZeneca's pipeline, ensures novel candidates are evaluated against a meaningful reference point.
Why Functional, Cell-Based Assays Matter Here
A recurring theme across the regenerative medicine and cancer literature is that gene expression and protein concentration alone are insufficient to characterize senescence's functional impact because what ultimately matters is whether a secreted factor is bioactive enough to trigger a response in a target cell population [2, 3, 4]. This is exactly the gap that validated, cell-based cytokine bioassays are designed to fill. A bioassay such as the IL-6/7TD1 proliferation assay shown above provides a direct, quantitative, and reproducible functional readout that complements ELISA-based concentration measurements, SA-β-Gal staining, and transcriptomic profiling, giving researchers a more complete picture of whether the cytokine output of a senescent population is biologically meaningful.
For groups working across this spectrum, whether characterizing SASP composition in an aging or wound-healing model, screening senolytic or senomorphic candidates, or validating pro-senescence cancer therapeutics, having access to a broad, pre-validated panel of cytokine and chemokine bioassays instead of building each one from scratch meaningfully shortens the path from hypothesis to data. It is an area where contract research support, including the kind of dose-response bioassay work and custom assay development illustrated here, can let academic and biotech teams focus their effort on the biological question rather than on assay re-development.
Looking Ahead
Senescence research is converging on a more nuanced model where context, duration, clearance, and SASP composition determine outcome rather than senescence being simply "good" or "bad" [3, 4, 9]. Single-cell and spatial multi-omic approaches are mapping senescent cell heterogeneity across tissues, while machine-learning classifiers are making senescence screening tractable even in cancer lines that already express baseline senescence markers [10]. As these tools mature, functional cytokine bioassays remain the critical validation layer, translating molecular and morphological signals into the quantitative, biologically grounded readouts that drug discovery and regenerative medicine programs need to move candidates forward.
At SBH Sciences, we apply our expertise in cell-based assays and cytokine profiling to support research into complex disease biology, including senescence and cancer. Our functional cellular models and multiplex cytokine platforms enable the study of senescence-associated phenotypes, tumor microenvironment signaling, and inflammatory pathways that can influence cancer progression and therapeutic response.
See how our cell-based assays can help you explore senescence as a potential therapeutic target in cancer research and drug discovery. Ready to move your research forward? Partner with SBH Sciences to generate the functional data needed to investigate senescence biology and support your cancer drug discovery program. Please contact us, we would be delighted to work with you!